(3)SIB1 株由来アルカリホスファターゼの低温適応に関する研究(2006年以後中断)
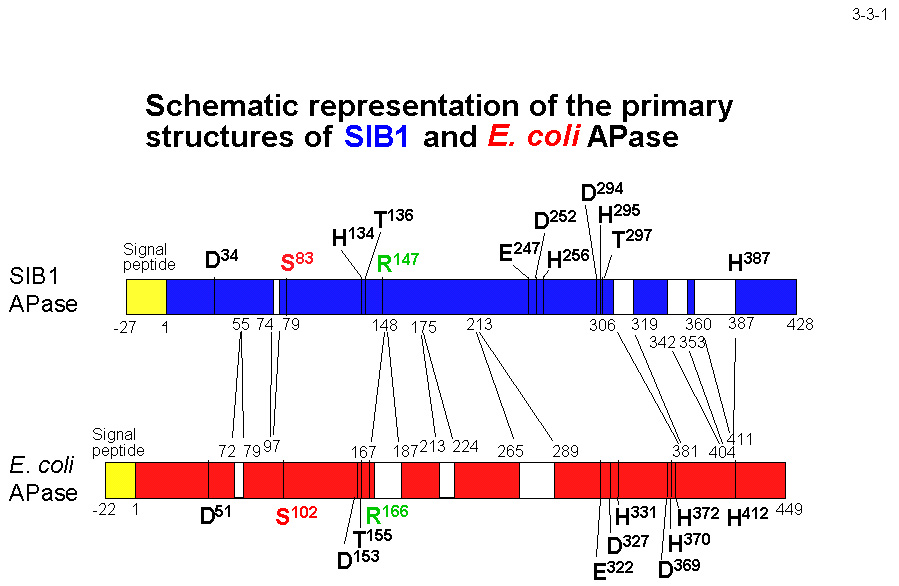
SIB1 株由来アルカリホスファターゼ(SIB1 APase)は生物界に広く存在するリン酸モノエステラーゼの一種で、455 アミノ酸残基から成る前駆体として合成された後ペリプラズムに分泌される(3-3-1)。 この際、27 残基から成るシグナルペプチドは N 末端から除去される。大腸菌APase同様ホモダイマーとして存在する(各サブユニットは 428 残基から成る)。SIB1 APase のアミノ酸配列はS. oneidensis
MR-1 株由来 APase (MR-1 APase)と最も高い同一性(66%)を示す。また、低温菌Shewanella sp. AP1 株由来 APase
(SCAPase)と 62%、低温菌 TAB5 株由来 APase
(TAB5 APase)と 55%、大腸菌 APase と 37%、
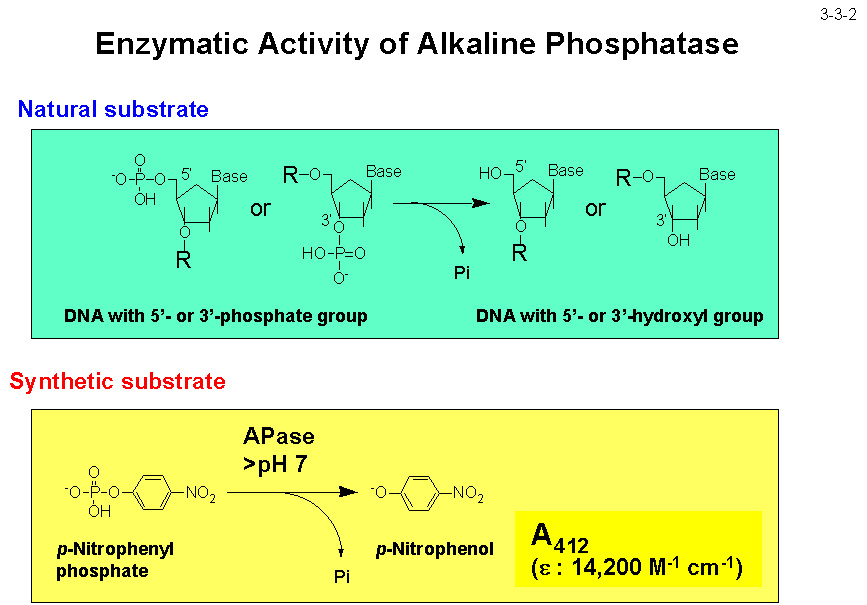
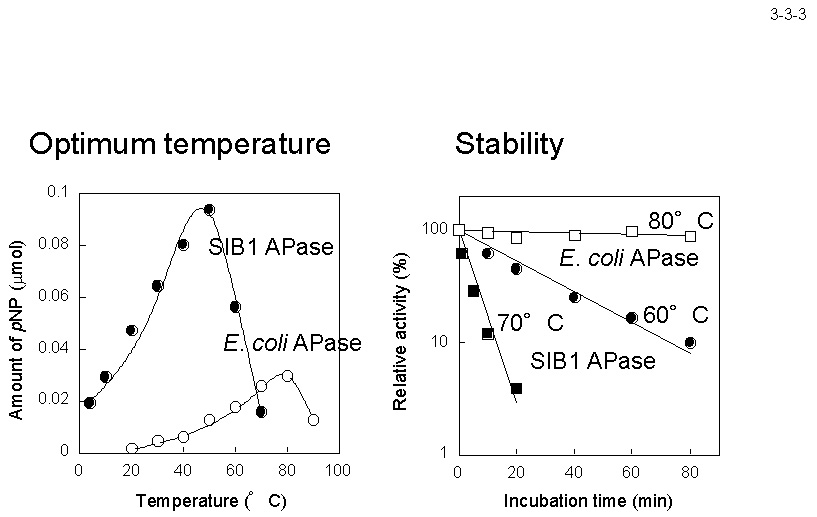
枯草菌 APaseIII と 42%、 Thermus sp. FD3041 由来 APase と 35%、高度好熱菌Thermotoga maritime由来 APase と 38% の同一性を示す。 本酵素の活性は p-ニトロフェニルリン酸を加水分解した時に生じる p-ニトロフェノールを 410 nm の波長で比色定量することにより簡単に測定できる(3-3-2)。SIB1 APaseの活性の至適温度は 50℃ で、 大腸菌 APase より 30℃ も低い(3-3-3)[Suzuki, Y. et al. (2005) Biosci. Biotechnol. Biochem.
69, 364-373]。両酵素の至適温度における活性を比較すると、SIB1 APase の活性は大腸菌 APase の活性より3ー4倍高い。また SIB1 APase は 70℃ では 3.9 分の半減期で失活し、 80℃ でも半減期が 6 時間以上もある大腸菌 APase に比べると著しく不安定である。従って、SIB1 APase は典型的な低温適応酵素の一つといえる。
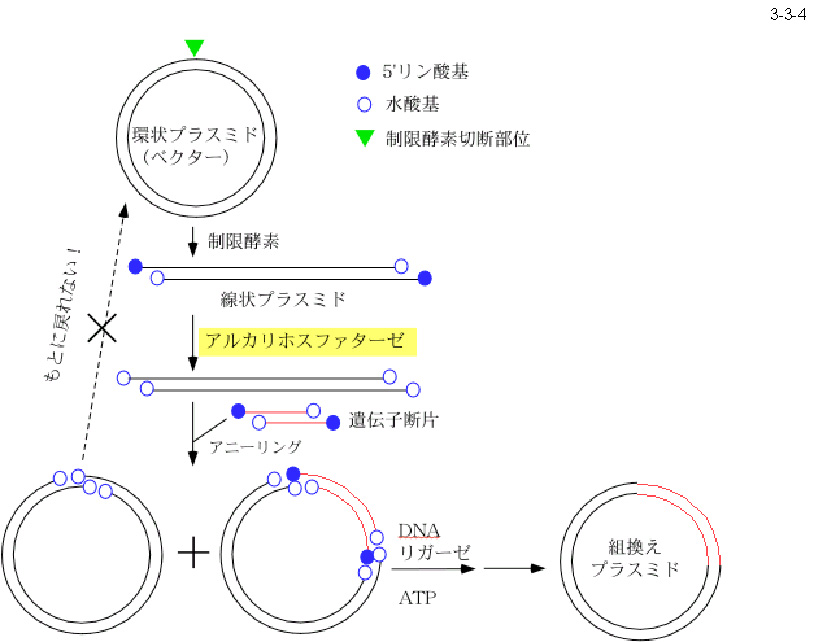
ところで、大腸菌 APase は BAP と呼ばれ、遺伝子工学実験において、DNA の 5’末端のリン酸基を除去する反応に用いられている(3-3-4)。しかし、 反応後の DNA はほとんどの場合 DNA 連結反応に用いられるが、APase はその反応を阻害してしまう。そのため、反応後の DNA 溶液に含まれている APase は完全に取り除くか、失活させなくてはならない。
BAP は熱に安定なので、フェノール/クロロホルム抽出など煩雑な作業によりその活性を除去する必要がある。しかし、熱に不安定な SIB1 APase を用いれば、加熱処理だけでその活性を除去できる。また、 SIB1 APase を用いれば 5’末端突出 2 本鎖 DNA の 5’末端リン酸基だけでなく 3’末端突出 2 本鎖 DNA や平滑末端 2 本鎖 DNA の 5’末端リン酸基も効率的に除去できる。
従って、SIB1 APase は DNA の 5’末端のリン酸基を除去するための遺伝子工学用試薬として BAP より有用と考えられる。現在、SIB1 APase は PAP としてフナコシから市販されている。
SIB1 APase と大腸菌 APase のアミノ酸配列を比較すると、SIB1 APase にはジスルフィド結合がなく(大腸菌 APase には2つある)、荷電アミノ酸残基も少ない。また、その配列には多くの欠失や挿入がみられる。
しかし、その他の APase のアミノ酸配列も比較してみると、荷電アミノ酸含量の減少、ジスルフィド結合数の減少、挿入配列の有無は必ずしも全ての低温菌由来 APase に当てはまる特徴ではなく、また低温菌 APase だけの特徴でもない。 TAB5 APase の荷電アミノ酸残基含量は 20.7% と大腸菌 APase と変わらず、中温菌由来の MR-1 APase、枯草菌 APaseIII はジスルフィド結合を有していない。TAB5 APase はその代わり Pro 残基の含量が 2.3% と大腸菌 APase や SIB1
APase より低くなっている。 これらの結果は、低温菌由来 APase がそれぞれ異なった戦略で不安定化していることを示唆している。SIB1 APase の低温適応機構を明らかにするためには、その結晶構造を決定し、中温菌由来APaseの活性や安定性との違いをもたらす因子を同定する必要がある。
なお、本研究は2006年以後中断している。
{kind=link}
{kind=link}
{kind=link}
{kind=link}