�i�Q�jSIB1�AMR-1 ���R�� RNase H �̒ቷ�K���Ɋւ��錤��
SIB1���́A�咰�ۓ��l type 1�iSIB1 RNase
H1�j�� type 2 (SIB1 RNase
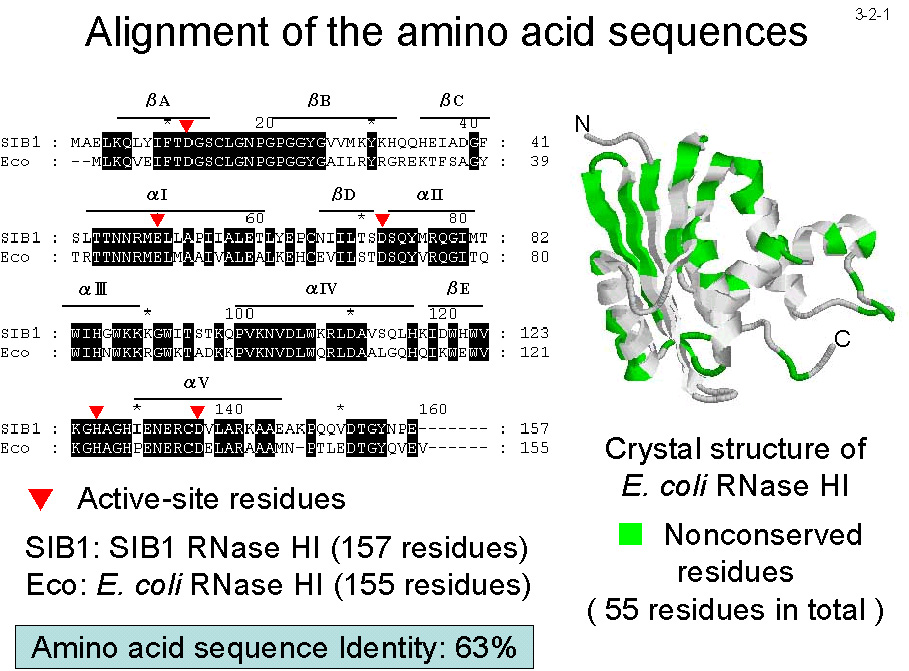
H2�j�̂Q��ނ� RNase H �����BSIB1 RNase H1�� 157 �A�~�m�_�c����Ȃ�A
�咰�� RNase H1�iEc-RNase H1�j�� 63% �̃A�~�m�_�z��̓��ꐫ������[Ohtani, N. et al.
(2001) Protein Eng. 14, 975-982]�B����ASIB1 RNase H2 �� 212 �A�~�m�_�c����Ȃ�A�咰�� RNase H2�iEc-RNase H2�j�� 64% �̃A�~�m�_�z��̓��ꐫ������[Chon, H. et al. (2006) FEBS J. 273, 2264-2275]�B ����������m�}�[�Ƃ��đ��݂���BSIB1 RNase H1 �� SIB1 RNase H2 �̂Q�������C�I���v�����A���K pH�A��ؒf���ʂȂǂ̍y�f�I�����͂��ꂼ�� Ec-RNase H1�AEc-RNase H2 �ɗގ����Ă���B
�܂�ASIB1 RNase H1 �� 5 mM Mg2+���݉��ōő劈���������̂ɑ��āASIB1 RNase H2 ��5 mM Mn2+���݉��ōő劈���������B�܂��ASIB1 RNase H1 �̎��K pH �� 8 �t�߂ł���̂ɑ��āA SIB1 RNase H2 �̎��K pH �� 10 �t�߂ł���B����ɁASIB1 RNase H1 �� Ec-RNase H1�ƁASIB1 RNase H2 �� Ec-RNase H2 �ƁA ���ꂼ��قړ����ʒu�� 12 mer RNA/DNA hybrid ����� 29 mer DNA13-RNA4-DNA12/DNA��ؒf����B
RNase H1�� RNase H2 �̑��ΓI�Ȋ�������萫�́ASIB1 �y�f�Ƒ咰�ۍy�f�ł͈ȉ��̂悤�ɑ傫���قȂ�B�P�jSIB1 RNase H1 �̔䊈���i2.2 u/mg�j�� SIB1 RNase H2 �̔䊈���i26 u/mg�j��蒘�����Ⴂ���A Ec-RNase
H1 �̔䊈���i9.5 u/mg�j�� Ec-RNase H2 �̔䊈���i1.8 u/mg�j��荂���B�܂�ASIB1 ���ɂ����Ă� RNase
H2 �̕��������͍������咰�ۂɂ����Ă� RNase
H1 �̕��������B �Q�jSIB1 RNase H1�̎��K���x�i30���j�� SIB1 RNase
H2 ���� 10�� �Ⴂ���AEc-RNase H1 �̎��K���x�i50���j�� Ec-RNase
H2���� 5�� �����B�܂��A�M������̎c���������r����� �i30���ɂ����锼�����FSIB1 RNase
H1�A1.5 min�GSIB1 RNase H2�A45 min�GEc-RNase H1�A��1 h�GEc-RNase H2�A60 min�j�ASIB1 RNase H1 �� SIB1 RNase H2 ��蒘�����s����ł��邪�AEc-RNase H1�� Ec-RNase H2 ���ނ������ł���B �R�j�咰�ۂő�ʐ��Y������ƁASIB1 RNase H1 �� Ec-RNase H2 �̓C���N���[�W�����{�f�B���`�����₷���̂ɑ��āASIB1 RNase H2 �� Ec-RNase H1 �͉n���ł���B�ȏ�̌��ʂ́A��v�Ȋ�����S�� RNase H �������ɂ���ĈقȂ� �i�咰�ۂɂ����Ă� RNase H1�ASIB1 ���ɂ����Ă� RNase H2�j���Ƃ��������Ă��苻���[���B
SIB1 ���R�� RNase H �̒ቷ�K���@�\�Ɋւ��ẮA���萫�⎊�K���x���傫���قȂ� SIB1 RNase H1 �� Ec-RNase H1 ���y�A�Ƃ��ĉ�͂������߂Ă���B
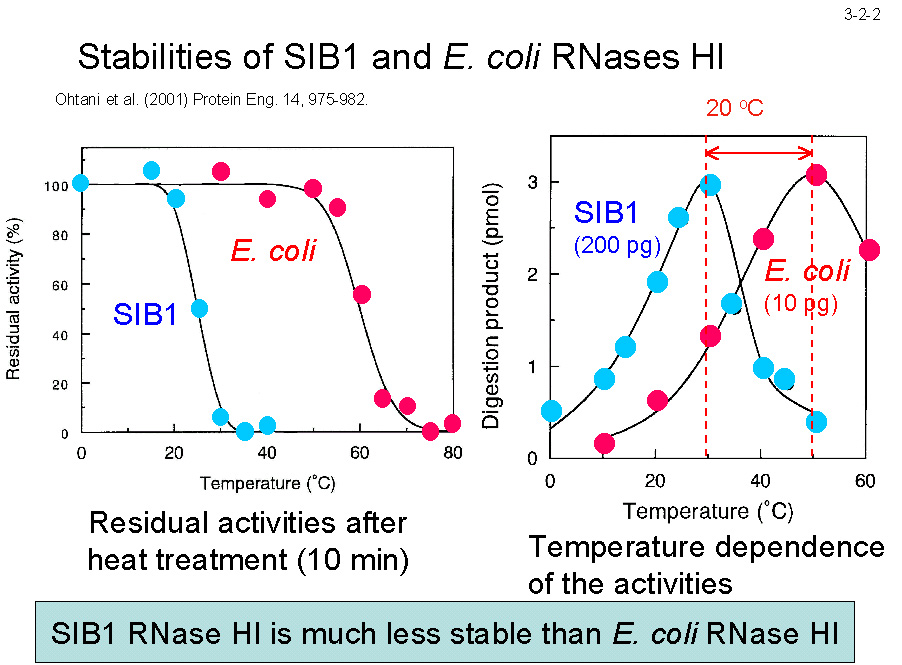
���҂̃A�~�m�_�z��̓��ꐫ�� 63% �Ɣ�r�I�����ɂ�������炸�i3-2-1�j�ASIB1 RNase H1 �� Ec-RNase H1 ��蒘�����s����ŁA���̊������K���x�� Ec-RNase H1 ���� 20�� �Ⴂ�i3-2-2�j�B
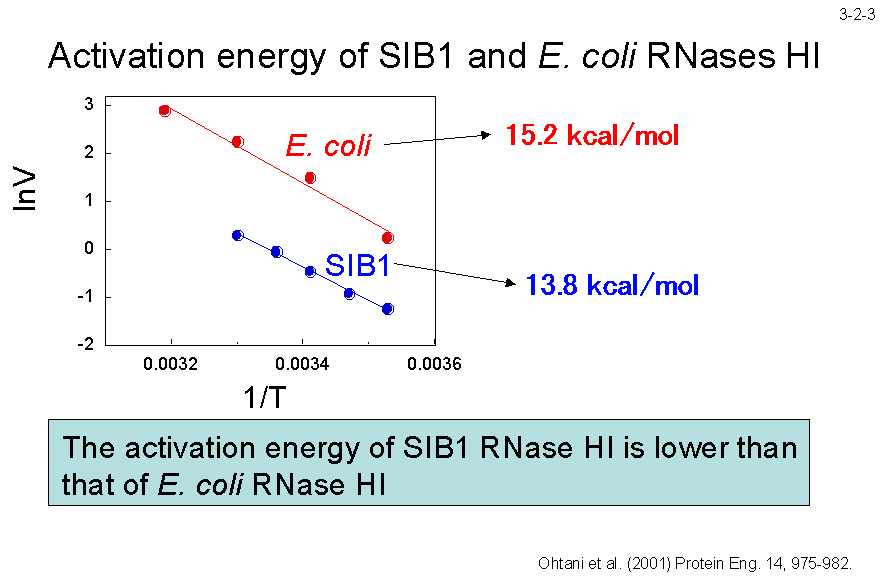
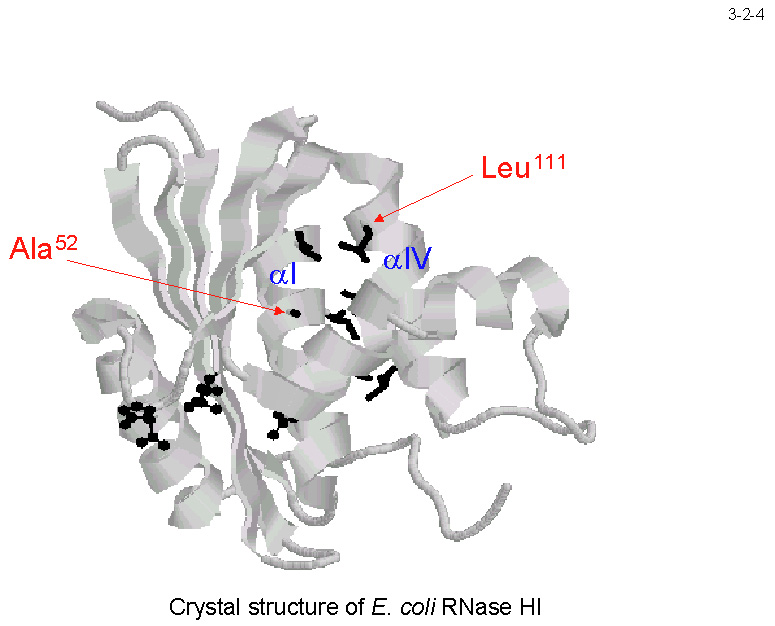
�܂��A�������G�l���M�[�� SIB1 RNase H1 �̕����Ⴂ�i3-2-3�j�B���҂̈ꎟ�\�����r����ƁA���w���b�N�XI�̒������̃A�~�m�_�� Ec-RNase H1 �ł� Ala�iAla52�j�ł���̂ɑ��� SIB1 RNase
H1 �ł� Pro�iPro54�j�ł���i3-2-4�j�B �܂��A���w���b�N�X �W �ɂ���a�����R�A���`������A�~�m�_�̈�� Ec-RNase H1 �ł� Leu�iLeu111�j�ł���̂ɑ���SIB1 RNase H1 �ł� Ser�iSer113�j�ł���i3-2-4�j�B
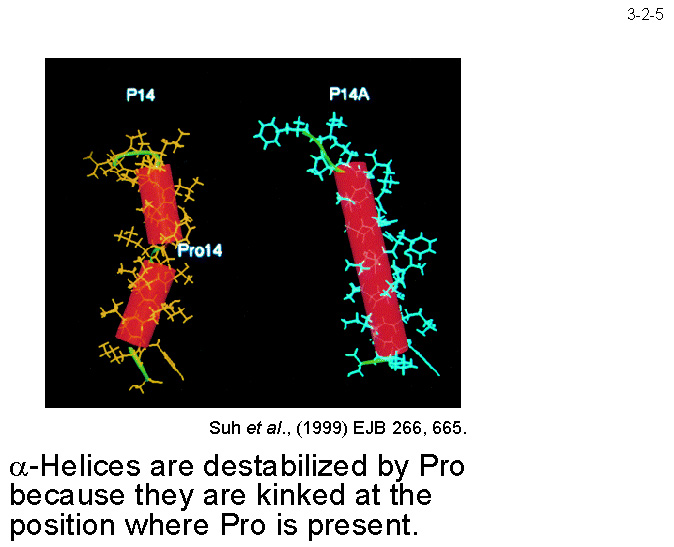
Pro �����w���b�N�X��s���艻���i3-2-5�j�ASer �͐e�����ł���̂ŁA�����̃A�~�m�_�u���� SIB1 RNase H1 �̒ቷ�K���Ɋ�^����ƍl������B���ہA
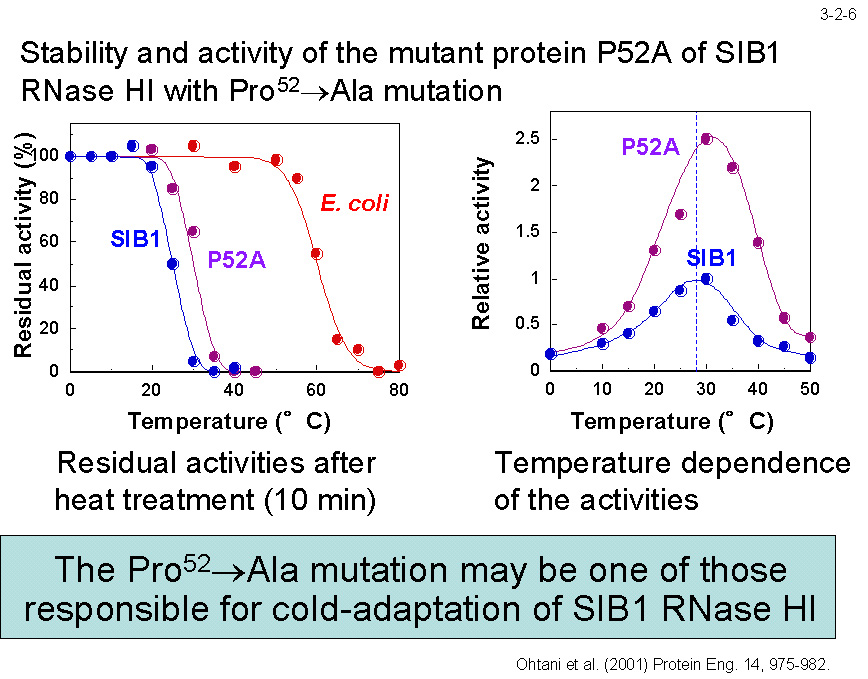
���ʓ��ٓI�ψٖ@�ɂ�� Ala52 �� Pro �ɒu������� Ec-RNase H1 �͖� 5�� �s���艻���i�����͕ω��Ȃ��j�A���� Pro55 �� Ala �ɒu������� SIB1 RNase H1 �͂����炩���艻����i����������j�i3-2-6�j[Ohtani,
N. et al. (2001) Protein Eng. 14, 975-982]�B
�������ASer113��Leu �ψقɂ�� SIB1 RNase H1 �͂ނ���s���艻����i�������ቺ�j�B�����炭���̃A�~�m�_�u�����������Ă��p�b�L���O�ɕs��������邽�� SIB1 RNase H1 �͈��艻���Ȃ����̂Ǝv����B
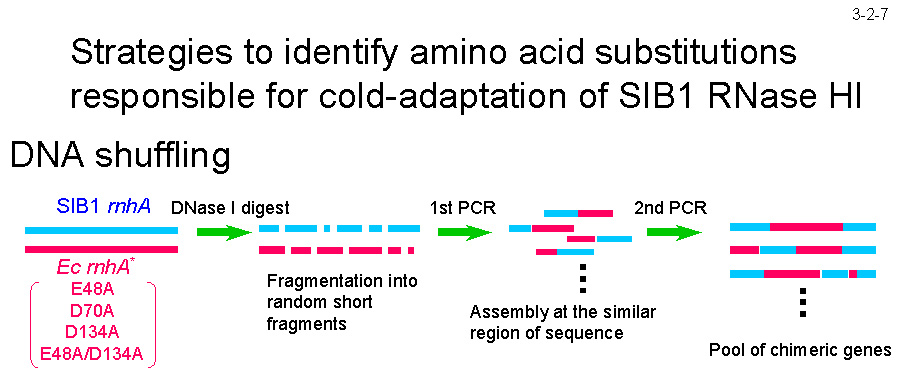
SIB1 RNase H1�� Ec-RNase H1 �̈��萫�̈Ⴂ�������炷�A�~�m�_�u���肷�邽�߂� DNA �V���t�����O�@�����݂Ă���i3-2-7�j�B DNA �V���t�����O�@�̓L������`�q��ׂɍ\�z������@�Ƃ��ėL���ł��邪�ASIB1 RNase H1 ������ȍy�f���R�[�h����L������`�q�iSIB1 RNase H1 ��`�q�� Ec-RNase H1 ��`�q�̃L������`�q�j�́A
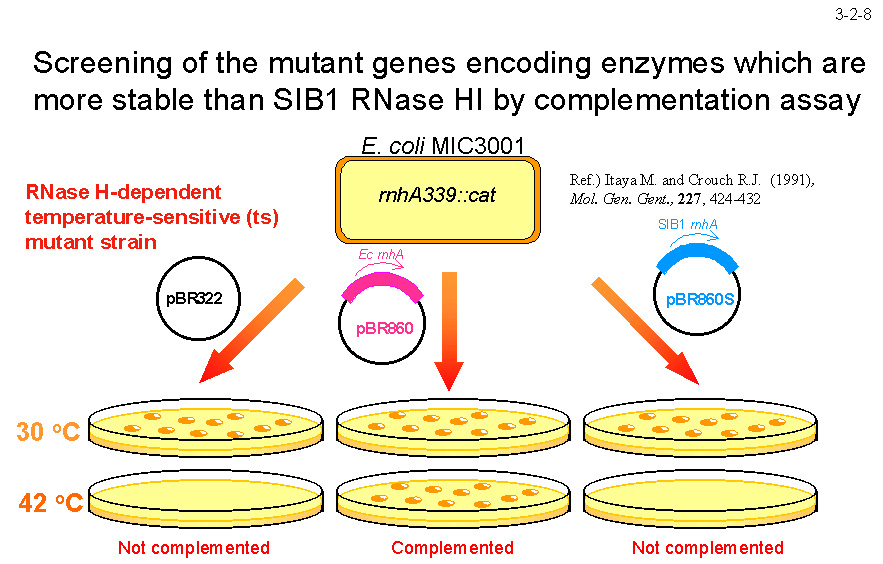
�咰�ەψي� MIC3001 �܂��� MIC2067��p���ăX�N���[�j���O����i3-2-8�j�B�����̑咰�ەψي��� RNase H1 ��`�q��j��Ă��邽�߂ɉ��x���̐����������B
�܂�AMIC3001 �� MIC2067 �� 42�� �ł͐��炵�Ȃ����AEc-RNase H1��`�q������� 42�� �Ő��炷��悤�ɂȂ�B������ SIB1 RNase H1 �� 42�� �ŔM�ϐ�����̂ŁA SIB1 RNase H1 ��`�q�����Ă� MIC3001 �� MIC2067 �� 42�� �Ő���ł��Ȃ��B�]���� MIC3001 ���邢�� MIC2067 �� 42�� �Ő���ł���悤�ɂȂ�L������`�q�� SIB1 RNase H1 ������ȃL�����y�f���R�[�h���邱�Ƃ����҂����B�������A����܂ł̂Ƃ��� Ec-RNase H1 �����������ȏ���߂�L�����y�f���������Ă��炸�ASIB1 RNase H1 �����艻����A�~�m�_�u���̓���ɂ͎����Ă��Ȃ��B
��q�� SIB1 RNase H1 �� P52A �ψّ̂��R�[�h�����`�q�� MIC3001 �̉��x���𑊕₷��̂ŁA���̂悤�ȕψّ̂��R�[�h����L������`�q�������Ȃ��̂́A DNA �V���b�t�����O�@�ɂ��L������`�q���C�u�����[�����҂ǂ��薳��ׂɍ\�z����Ă��Ȃ����߂ƍl������B�����Ō��݁ASIB1 RNase HI �Ƃ̑���������荂��Shewanella oneidensis
MR-1 ���R�� RNase H1 �iMR-1 RNase H1�j���p�[�g�i�[�Ƃ���L�����y�f�̍\�z�����݂Ă���B
���Q�O�O�V�N�Ȍ�̌�����
�P�jSIB1������m���[��hybrid binding doamin�iHBD�j��L����V�Ktype 1 RNase H�iSIB1 HBD-RNase H1�j��P��������������͂����B���̌��ʁA�{�y�f��SIB1-RNase H1�Ɠ��l�̂Q�������C�I���ˑ������������ƁA���̊�����SIB1-RNase H1����T�{�������ƁA�m���[��HBD�͊�����ɏd�v�ł��邱�Ƃ𖾂炩�ɂ���[Tadokoro, T. et al. (2007) FEBS J. 274, 3715-3727]�B
�Q�jMR-1 RNase H1�͂P�T�W�A�~�m�_�c��琬��A�咰��RNase H1�ƂU�V���̃A�~�m�_���̓��ꐫ�������BMR-1 RNase H1�̌����\�������肷��ƂƂ��ɁA���̊�������萫��咰��RNase H1�Ɣ�r�����B���̌��ʁA���q�����̃C�I���y�A�̐������Ȃ����Ƃƕ��q�\�ʂ̋ɐ��c��̊��������Ȃ����Ƃ�������MR-1 RNase H1�̍\���͑咰��RNase H1�Ƃقړ����ł��邱�ƁAMR-1 RNase H1�̕����咰��RNase H1����22���s����ł��邱�Ƃ𖾂炩�ɂ���[Tadokoro, T. et al. (2007)
Biochemistry 46, 7460-7468]�B�܂��A���ʓ��ٓI�ψٖ@�ɂ��A���q�����̃C�I���y�A�̌�����MR-1 RNase H1�̕s���艻������ł��邱�Ƃ𖾂炩�ɂ����B
�R�jMR-1 RNase H1��MIC3001�̉��x���𑊕�ł��邪�A���̂b���[���T�c�������ƕs���艻���邽�ߑ���ł��Ȃ��Ȃ�B���̕s���艻����MR-1 RNase H1�ψّ̂Ƀ����_���ψق����A���萫�̉����ψّ̂��X�N���[�j���O���邱�Ƃɂ��A�S�̈��艻�ψق肷�邱�Ƃɐ�������[Tadokoro, T. et al. (2008) Biochemistry 47, 8040-8047]�B�T�v���b�T�[�ψٖ@�ɂ�蓯�肳�ꂽ�����̈��艻�ψق�g�ݍ��킹�邱�Ƃɂ��MR-1 RNase H1�̈��萫�͖�19�����サ���B���̌��ʂ́A�Ǐ��I�ȍ\���̌��ׂ��C�����邱�Ƃɂ��ቷ�ۗR���y�f�̈��萫�𒆉��ۗR���y�f�̃��x���܂ō��߂邱�Ƃł��邱�Ƃ��������Ă���B.
�S�j�T�v���b�T�[�ψٖ@�ɂ�蓯�肳�ꂽ�S��ނ̈��艻�ψقƕ��ʓ��ٓI�ψٖ@�ɂ�蓯�肳�ꂽ�Q��ނ̈��艻�ψق�g�ݍ��킹�邱�Ƃɂ���29�����艻����MR-1 RNase H1�ψّ̂̌����\�������肷�邱�Ƃɂ��AMR-1 RNase H1�͋Ǐ��I�ȍ\���̌��ׂ̑g�ݍ��킹�ɂ��s���艻���Ă��邱�Ƃ𖾂炩�ɂ���[Rohman, M. S. et al. (2009) FEBS J. 276, 603-613]�B
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}