(1)SIB1 FKBP22 の構造と機能の研究
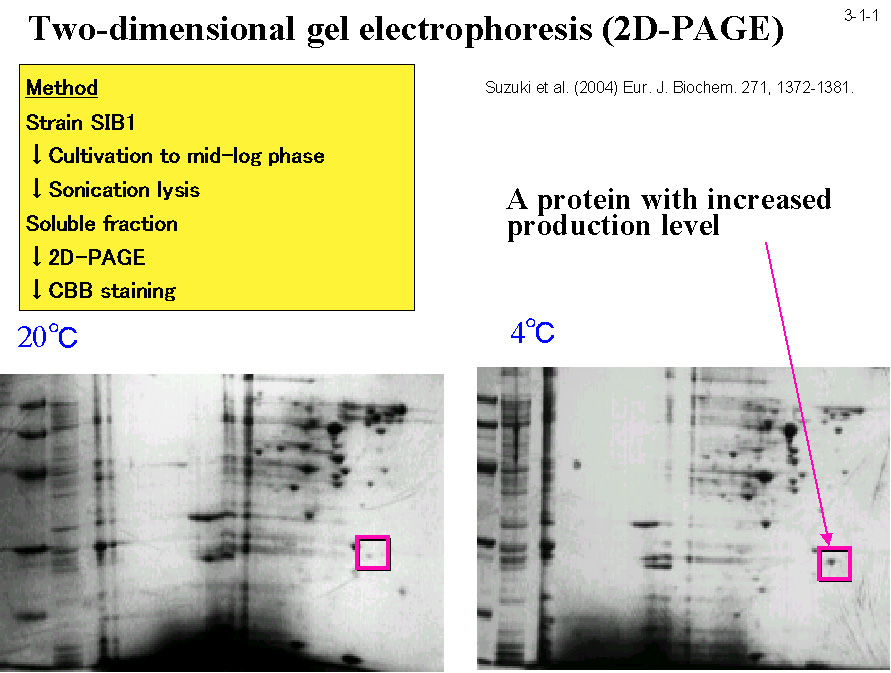
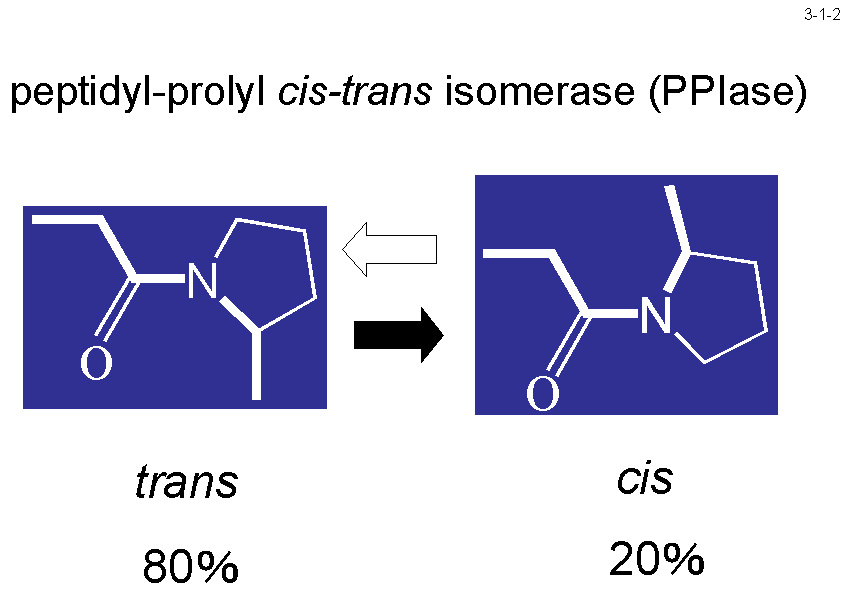
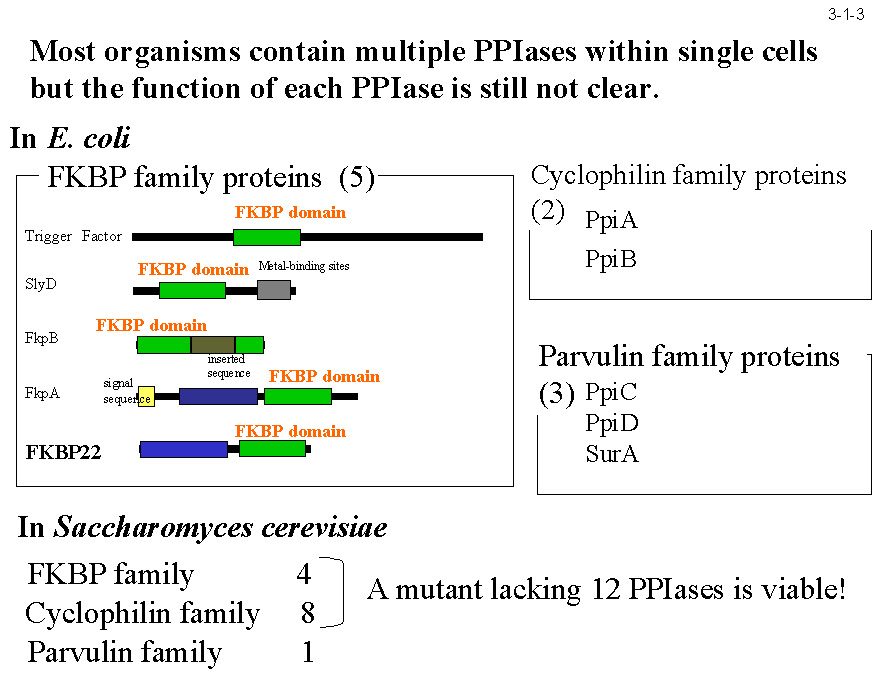
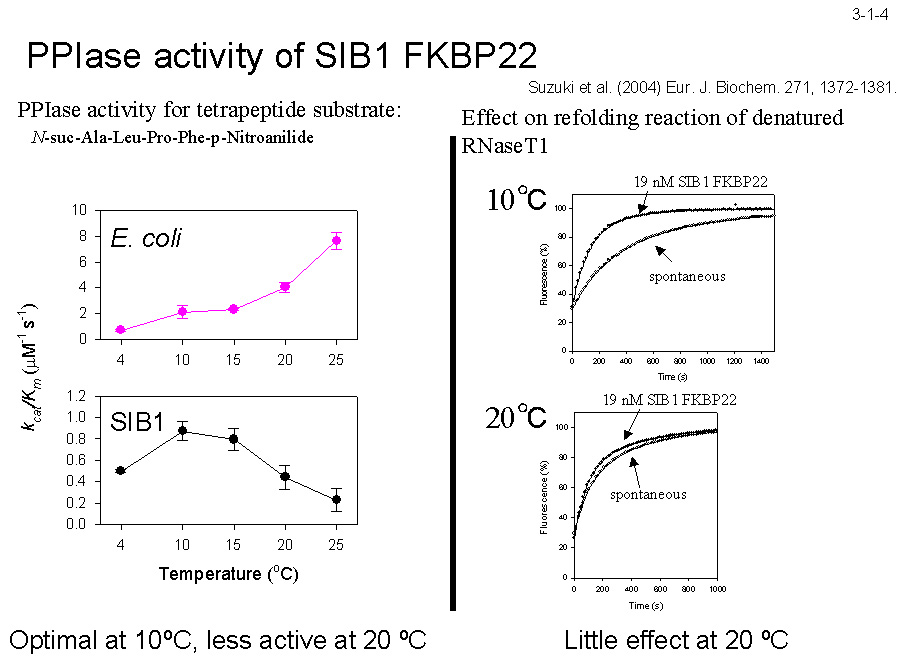
低温菌Shewanella sp. SIB1 株を 4℃ と 20℃ で生育させた時に菌体内に蓄積する蛋白質を二次元電気泳動で比較すると、いくつかの蛋白質が 4℃ で生育させた時に特異的に蓄積する。 これらの蛋白質は本菌の低温適応に関与すると考えられる。私達はそのうちの一つが大腸菌 FKBP22 と 56%、ヒト FKBP12 と 43%、L. pneumophila MIP と 41% のアミノ酸配列同一性のある蛋白質であることを見いだし(3-1-1)、 この蛋白質(SIB1 FKBP22)の構造や機能の研究を行っている。SIB1 FKBP22 は 205 アミノ酸残基から成るが、ペプチジルプロリルシストランスイソメラーゼ(PPIase)の3つのファミリーの一つである FKBP ファミリーに分類される(3-1-2)。 他の2つのファミリーはシクロフィリンとパーブリンである。PPIase 活性はプロリン残基N末端側にあるペプチド結合のシス−トランス異性化を触媒する(3-1-3)。SIB1 FKBP22 も PPIase 活性を示すが、 プロリン残基を含むテトラペプチドだけでなく、プロリン異性化反応が律速であることが知られている RNase T1 の折りたたみ反応も促進するので、その PPIase 活性は非特異的と考えられる(3-1-4)。 また、その活性は 10℃ で最大になり 20℃ 以上では急激に低下するので、本蛋白質は低温でのみよく働くと考えられる。プロリンの異性化反応は蛋白質折りたたみ反応の律速段階となることが知られており、その速度は低温では著しく低下するので、 SIB1 株は PPIase 活性をもつ FKBP22 の生産量を増やすことにより低温に適応していると考えられる[Suzuki, Y. et al. (2004) Eur. J. Biochem.
271, 1372-1381]。
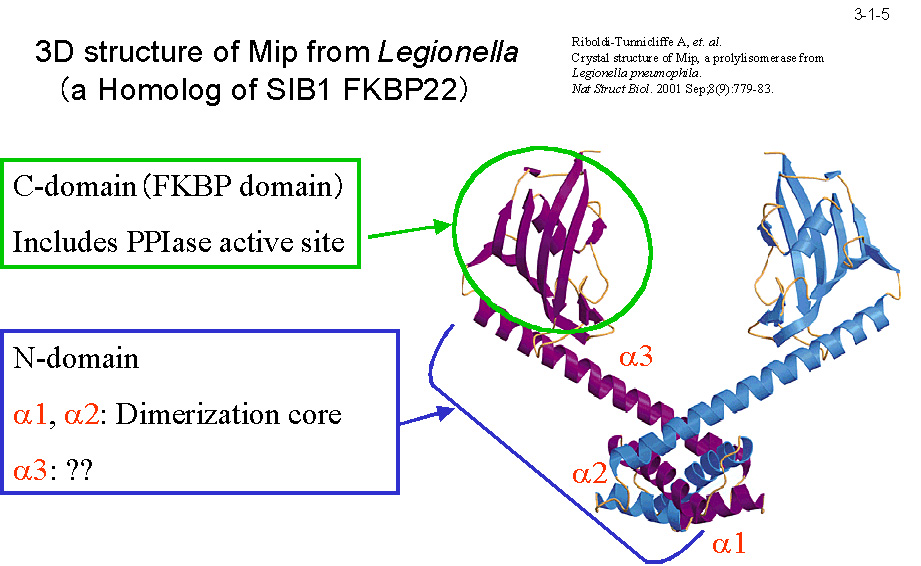
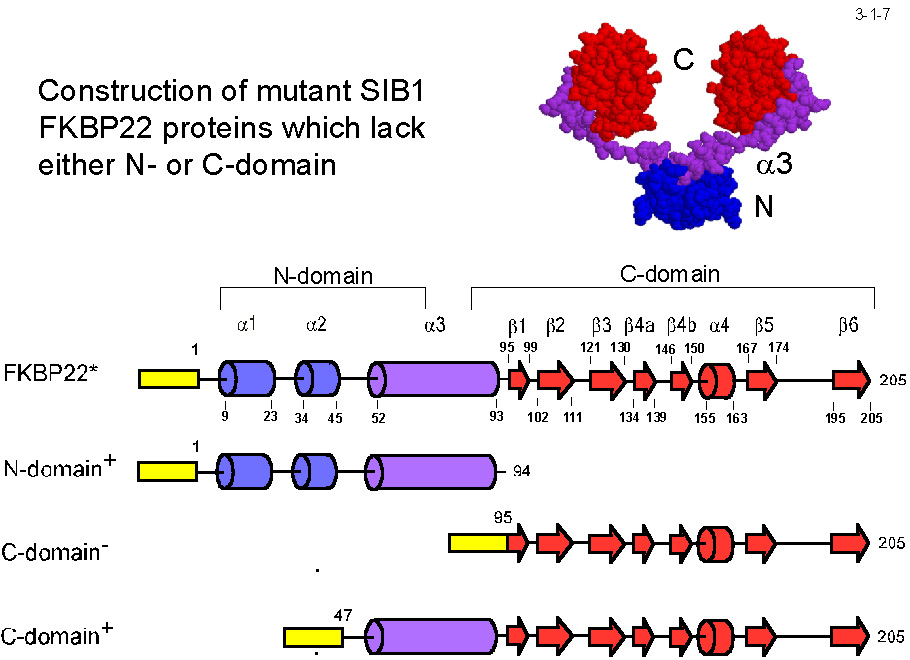
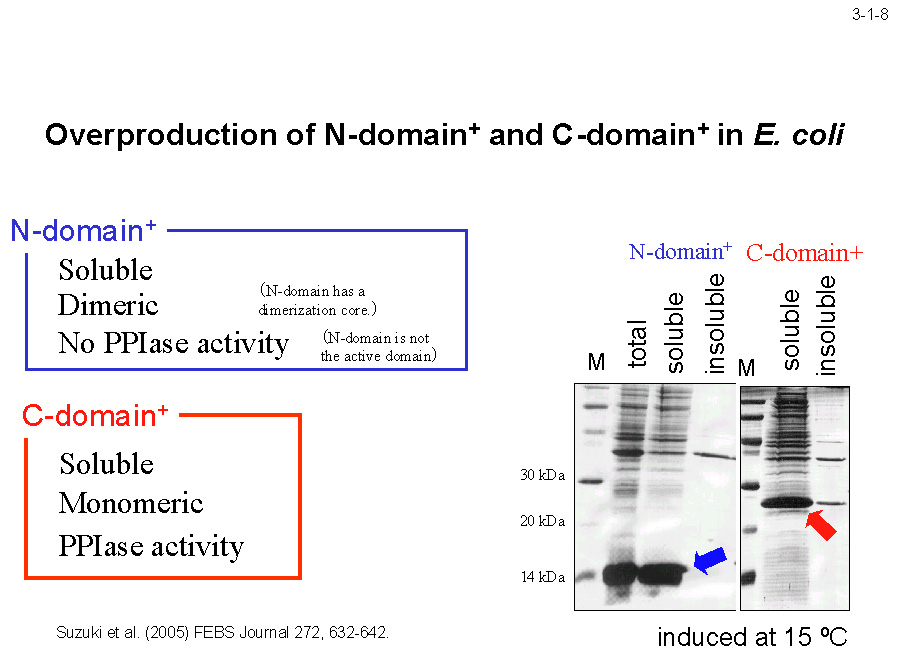
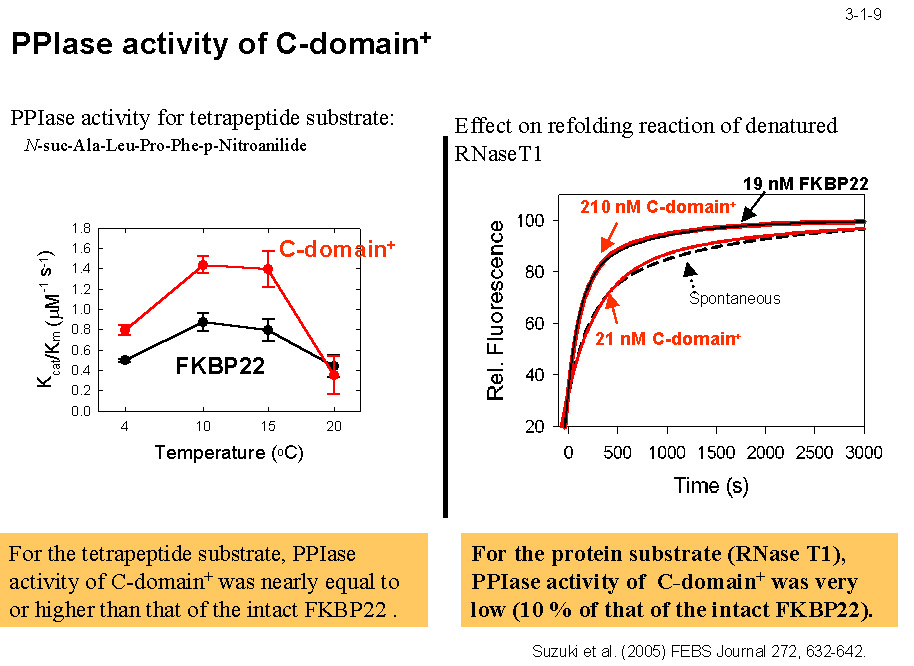
MIP の結晶構造(3-1-5)に基づいて構築した三次構造モデルによると、SIB1 FKBP22は PPIase 活性部位を含むC末ドメイン、二量化コアを形成する N 末ドメイン、 及び両者の間に位置する長いヘリックス(α3)からなる(3-1-6)。N 末ドメイン、C 末ドメインをそれぞれ N 末ドメイン+ α3(N-domain+)、C 末ドメイン+ α3(C-domain+)として発現させると(3-1-7) いずれも可溶性蛋白質として得られる(3-1-8)。N-domain+ は2量体として、C-domain+ は単量体として存在する。また、C-domain+ は PPIase 活性を示すが N-domain+ は活性を示さない。 ただし C-domain+が活性を示すのはテトラペプチドを基質とする時だけで、蛋白質(RNase T1)を基質とする時は活性を示さない(3-1-9)。SIB1
FKBP22 およびそのドメインと部分変性蛋白質(還元型αーラクトアルブミン) の結合を BIA-core を用いて解析すると、SIB1 FKBP22 および N-domain+ は結合するが、C-domain+ は結合しない。また N-domain+ は未変性蛋白質(非還元型αーラクトアルブミン)とは結合しない。
従って、N 末ドメインは部分変性状態の蛋白質に結合するのに重要と考えられる[Suzuki, Y. et al. (2005) FEBS Lett. 579, 5781-5784]。
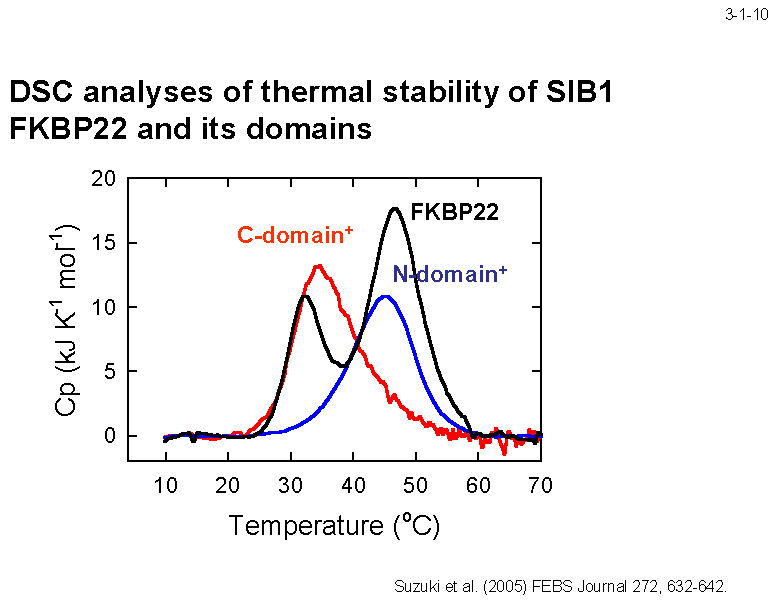
SIB1 FKBP22(全長)の熱変性を DSC により解析すると 32.5℃ と 46.6℃の2つのピークが観察される(3-1-10)。一方、C-domain+ の場合は 35.6℃ に、 N-domain+ の場合は 44.7℃ にそれぞれピークが1つだけ観察される。従って、SIB1 FKBP22の熱変性の2つのピークは、低温側が C 末ドメイン、高温側が N 末ドメインの変性に対応すると考えられる。 SIB1
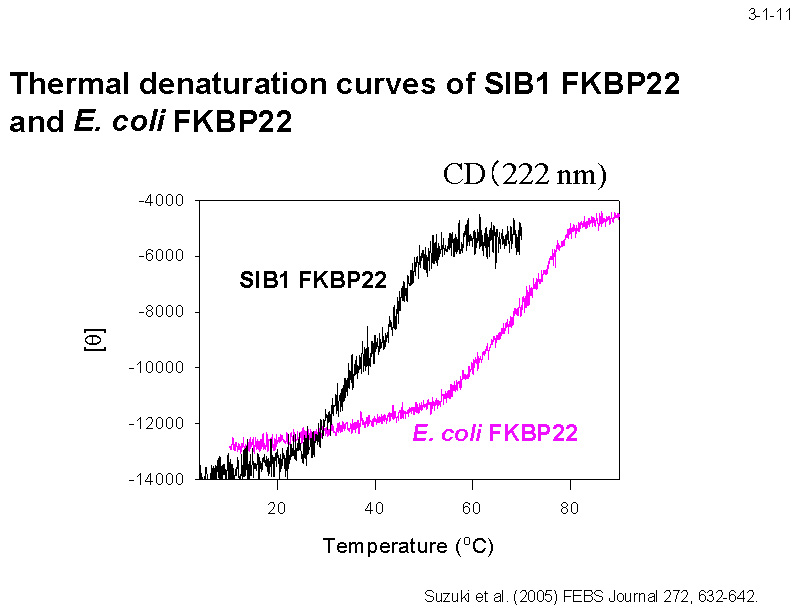
FKBP22 の熱変性を CD で解析しても2つに分離した転移が同様に観察される(3-1-11)。一方、大腸菌 FKBP22 の場合はこのように2つに明確に分離した転移は観察されず、 しかもその変性の転移温度は大きく(約 30℃)高温側にシフトしている。複数のドメインからなる低温適応酵素の中には、各ドメインの熱安定性が大きく異なる例が報告されており、低温での効率的な触媒活性に必要な柔らかさと基質の結合に必要な硬さを同時に実現するための戦略であるという提案がなされている。 SIB1 FKBP22 も同様の戦略で低温に適応しているのかもしれない[Suzuki,
Y. et al. (2005) FEBS J. 272, 632-6421]。
SIB1 FKBP22 はプロリン異性化反応を低温で触媒するので、シス型プロリンをもつために折り畳み反応が遅く凝集しやすい蛋白質の可溶化に利用できるかもしれない。
プロリンの異性化反応は、in vitro でのフォールディング実験においてしばしば律速段階となることが知られており、プロリン異性化酵素は in vivo での蛋白質折り畳み反応を促進すると考えられている。そこで、 凝集しやすい蛋白質を大腸菌で発現させる時に SIB1 FKBP22 を共発現させたり、尿素などで変性させた後で再構成する時に SIB1 FKBP22 を共存させたりする方法が、蛋白質の凝集を防ぐのに有効であるかもしれない。 蛋白質が凝集しやすい場合、通常、培養温度を下げたり発現誘導を抑制したりして蛋白質の合成速度を低下させることによりその凝集を防ぐ。しかし、温度を下げるとプロリンの異性化反応の速度も低下するので、この異性化反応が律速となる蛋白質の凝集を防ぐことができない。 SIB1 FKBP22 は低温に適応しているので、このような蛋白質を凝集させずに発現させるのに効果的であると期待される。
<2006年以後の研究>
1)SIB1 FKBP22のニ量体構造の役割:SIB1 FKBP22のニ量体構造の役割を明らかにする目的で、ニ量体を形成できない変異体NNC-FKBP22を構築した。この変異体は、N末端領域(Met8-Ala60)を重複させることによりN末端領域だけが野生型と同じニ量体構造を形成するように設計した。NNC-FKBP22の諸特性を解析することにより、NNC-FKBP22はモノマーとして存在すること、そのペプチド基質に対する活性はニ量体を形成する野生型体酵素よりむしろ向上すること、タンパク質基質に対する活性やシャペロン活性は野生型体酵素より大きく低下することを明らかにした [Budiman, C. et al. (2009) FEBS J. 276, 4091-4101]。以上の結果、ニ量体構造は部分変性状態のタンパク質基質への結合に必要であることを明らかにした。
2)SIB1 FKBP22のN末ドメインの結晶構造解析:SIB1 FKBP22のN末ドメイン(SN-FKBP22, residues 1-68)の結晶構造を決定した。その結果、N末ドメインだけでもニ量体を形成すること、全域にわたってニ量体の界面が形成されること、Val37とLeu41 のジッパー(Val-Leu knot)がコア領域を形成すること、これらのアミノ酸を両方ともArgに置換することによりこのジッパーを壊すと、変性してモノマーに解離することを明らかにした。以上の結果、 Val-Leu knot はSIB1 FKBP22のニ量体形成に不可欠であること、ニ量体化はSIB1 FKBP22のN末ドメインの構造形成に不可欠であることを明らかにした[Budiman, C. et al. (2011) Protein Sci. 20, 1755-1764]。
3)SIB1 FKBP22の活性部位残基の役割:SIB1 FKBP22の基質結合ポケットを構成する疎水性残基のうちのTrp157とPhe197、このポケットの近くに存在する極性残基のAsp137とArg142を他のアミノ酸残基に置換し、得られた変異体の活性、シャペロン活性を解析することにより、Trp157とPhe197は基質結合ポケットの形を最適な形に保つのに必要であり、Asp137とArg142は触媒活性に必要であることを明らかにした。また、活性部位の変異はシャペロン活性には影響を及ぼさないことを明らかにした。以上の結果に基づき、Asp137またはArg142がPro主鎖の窒素原子と水素結合することによりProのN末側のC−Nペプチド結合を回転させやすくするという触媒機構を提唱した[Budiman, C. et al. (2012) FEBS J. 279, 976–986]。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}