(1)リボヌクレアーゼ H(RNase H)の触媒機構と基質認識機構に関する研究
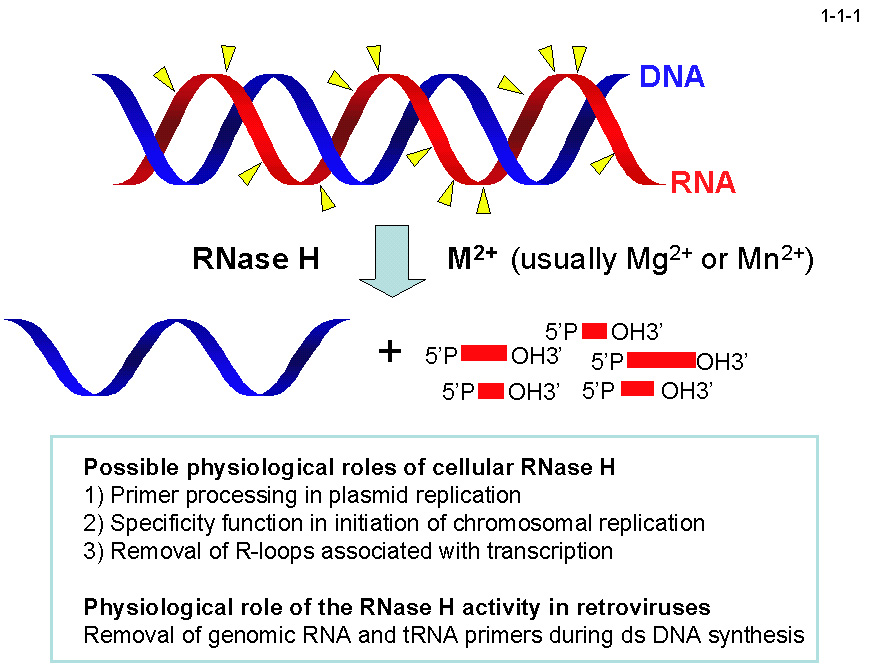
RNase H は RNA/DNA ハイブリッドの RNA鎖 のみを塩基非特異的に切断して、
5'位にリン酸基、3'位に水酸基をもつ RNA オリゴマーを生成するエンドリボヌクレアーゼで、生物界に普遍的に存在する(1-1-1)。
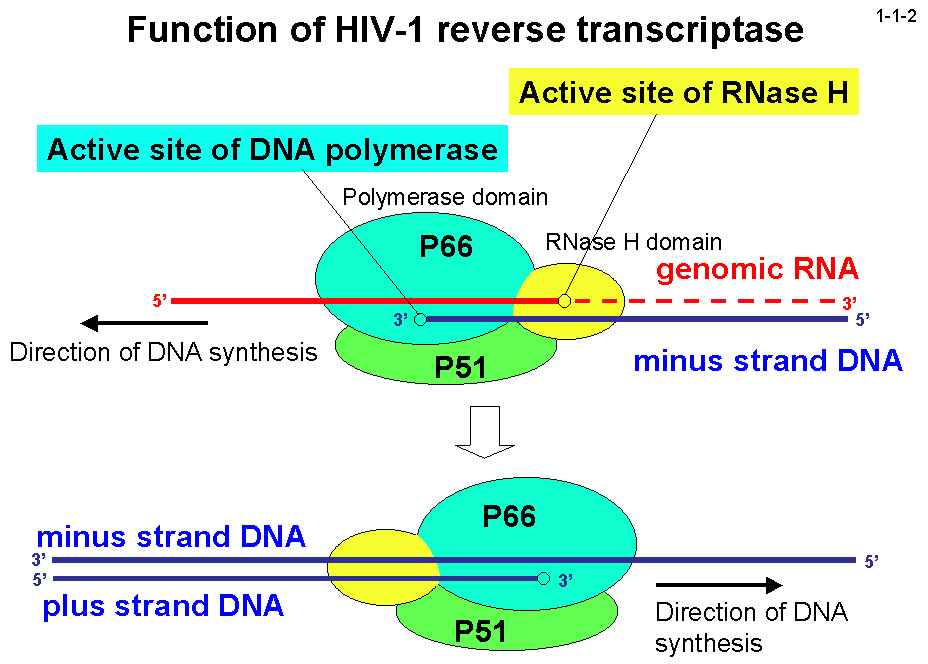
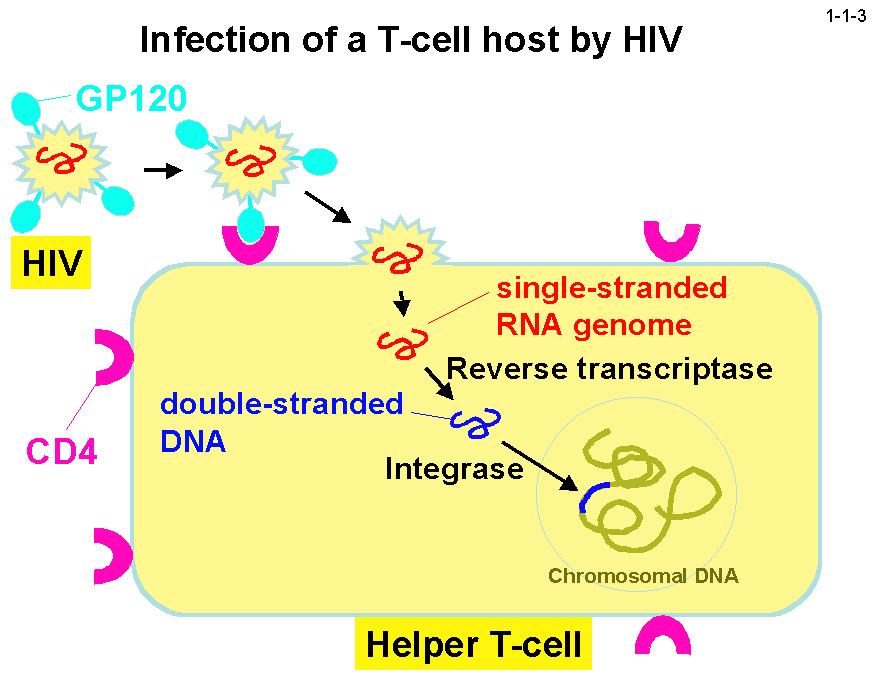
RNase H は逆転写酵素のC末端ドメインとしても存在するが(1-1-2)、その活性は HIV-1 の増殖に必須であるため(1-1-3)、
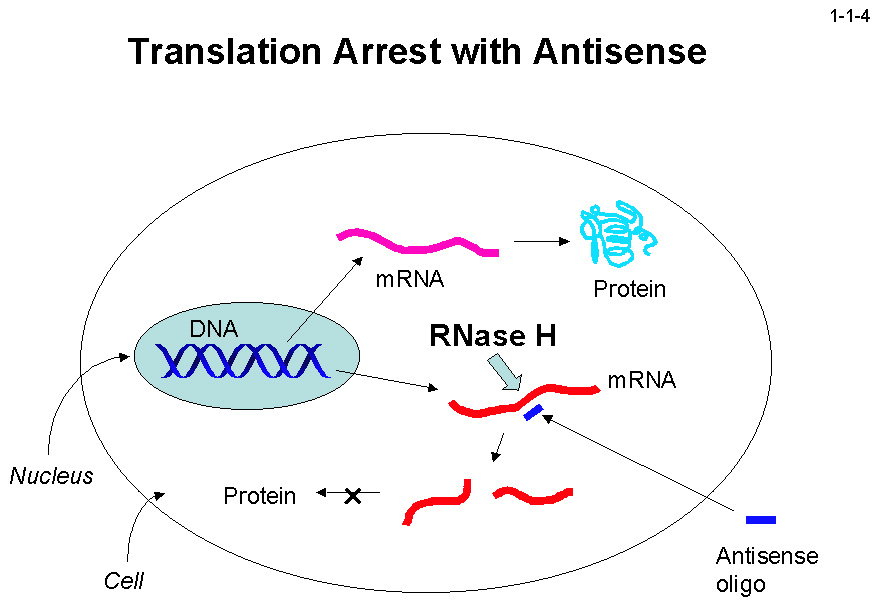
その阻害剤はエイズの治療薬になると期待されている。また、RNase H はアンチセンスオリゴヌクレオチドによる翻訳阻害にも深く関与する(1-1-4)。
従って、RNase H の生理機能を明らかにするためにも、新たなエイズ治療薬やアンチセンス医薬を開発するためにも、
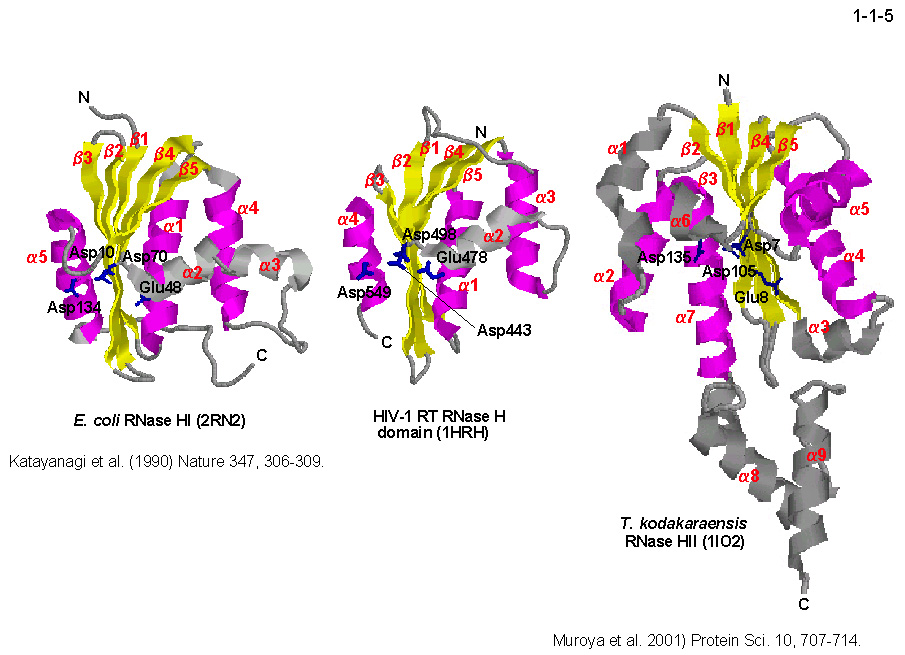
RNase H の触媒機構や基質認識機構に関する研究は重要である。これまでに大腸菌 RNase HI、HIV-1逆転写酵素、超好熱菌 RNase HII など
いくつかの RNase H の三次構造が決定されている(1-1-5)。これらの構造は RNase H-fold とよばれ、
ポリヌクレオチド基転移酵素の基本構造単位として生物界に普遍的に存在することが明らかにされている。
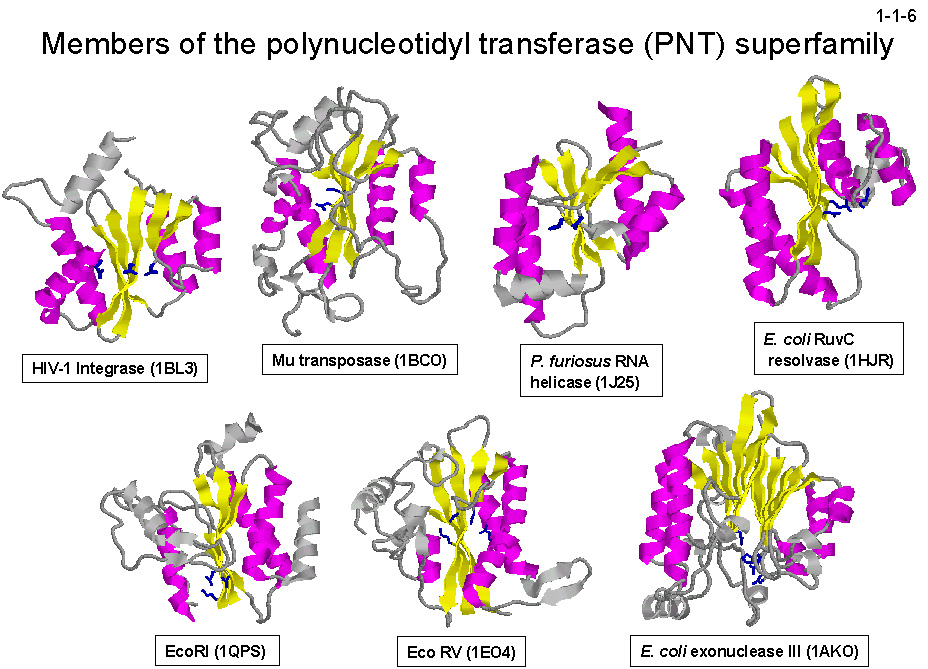
各種 RNase H はもちろんのこと、リゾルベース、インテグレース、トランスポゼース、RNA ヘリカーゼ、II 型 DNA 制限酵素など、
RNase H とは一次構造的にも機能的にも異なる酵素が三次構造的には RNase H-fold を持つ(1-1-6)。
これらの酵素に共通するのは、活性部位が複数の酸性残基で構成されること、そこに2価金属イオンが結合して活性型になること、
そして2本鎖核酸を認識し P-O3'結合を切断することである。従って、RNase H の触媒機構や基質認識機構を明らかにすることができれば、
同じスーパーファミリーに属する他の酵素の触媒機構や基質認識機構を理解するのにも役立つ。
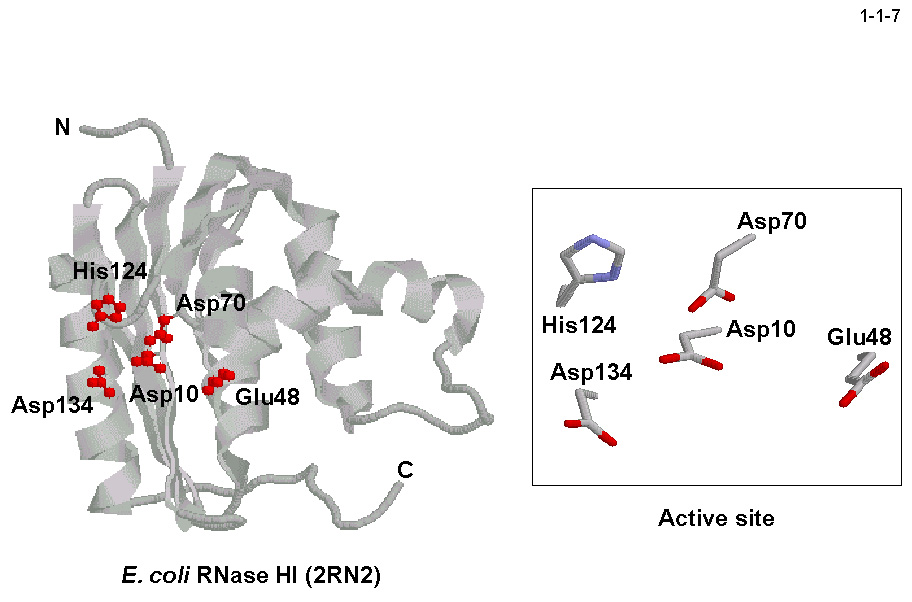
私達は、大腸菌 RNase HI をモデルとしてその触媒機構や基質認識機構を解析している。本酵素の活性部位は Asp10、Glu48、Asp70、Asp134 の
4残基の酸性アミノ酸と His124により構成される(1-1-7)。ちなみに type 2 RNase H の場合はその活性部位は4残基の酸性アミノ酸だけで構成される。
これらの酸性アミノ酸残基は2価金属イオンの結合や一般酸塩基として働く水分子の保持に必要で、ヒスチジン残基は一般塩基として働く水分子の活性化に必要と考えられている。
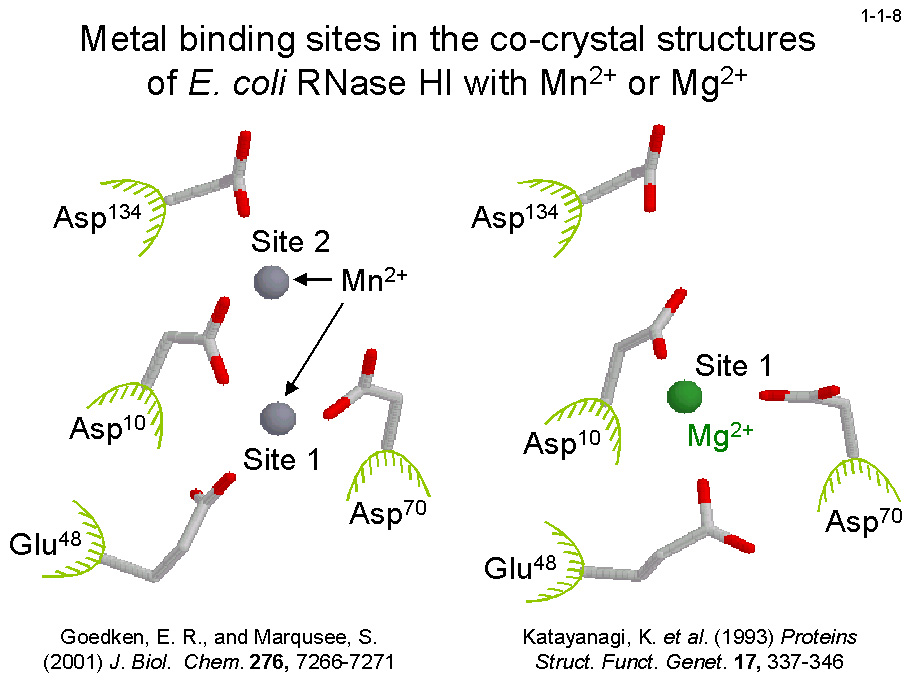
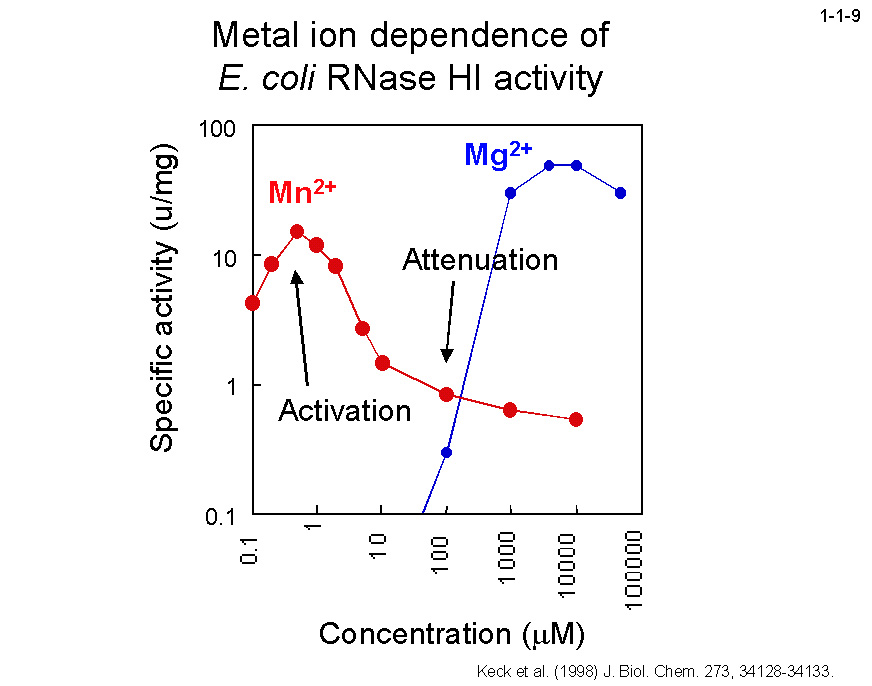
しかし、大腸菌RNase HIと2価金属イオンの共結晶構造によると、本酵素には Mg2+の場合は1原子、Mn2+の場合は2原子結合する(1-1-8)。
Mg2+結合部位は Mn2+結合部位(サイト1と2)のいずれとも一致しない。本酵素の至適Mn2+濃度が ~1 mM と極めて低いことから、1つめの Mn2+が結合すると酵素は活性型になり、
2つめの Mn2+が結合するとその活性は低下すると考えられている(1-1-9)。ちなみに、本酵素の至適 Mg2+濃度は数 mM と高い。
おそらく Mg2+は Mn2+より酵素に対する親和性が著しく小さいため高濃度にしても酵素に1原子しか結合しないと考えられる。
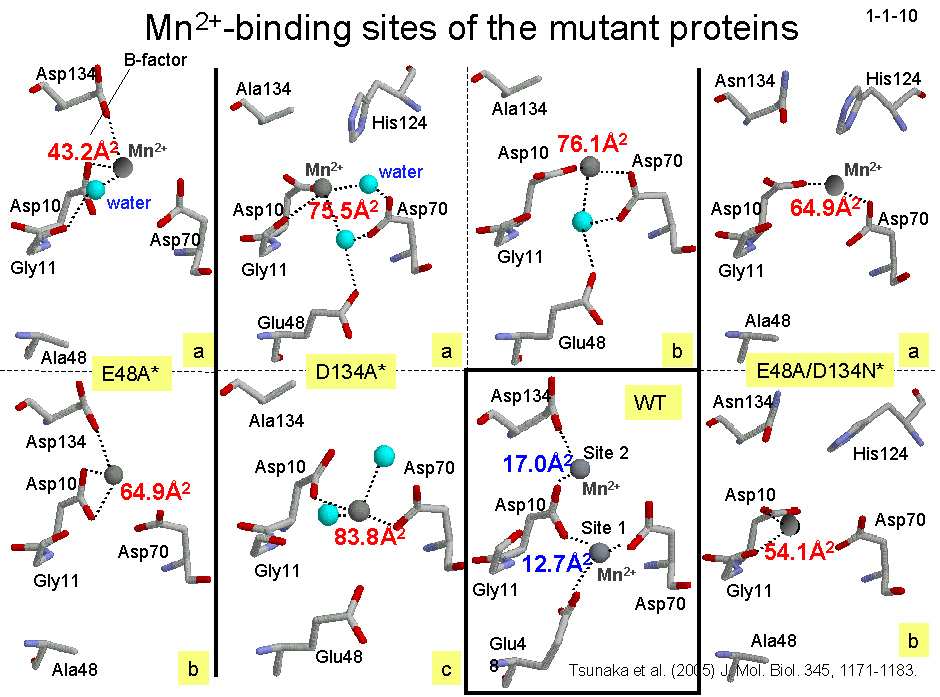
私達は、大腸菌 RNase HI の活性中心変異体と2価金属イオンの共結晶構造を解析することにより、1つめの Mn2+が結合するのはサイト1でも2でもなく
活性部位の中心付近であること、酵素が高い活性を示すためには活性部位に結合した2価金属イオンがゆらいでいる必要があること(金属イオンゆらぎモデル)などを提案している(1-1-10)。
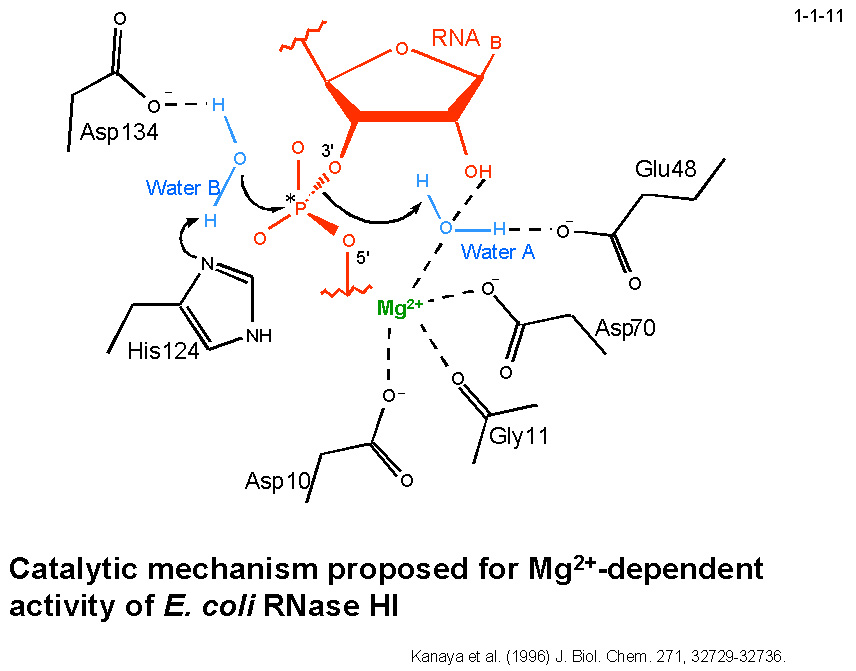
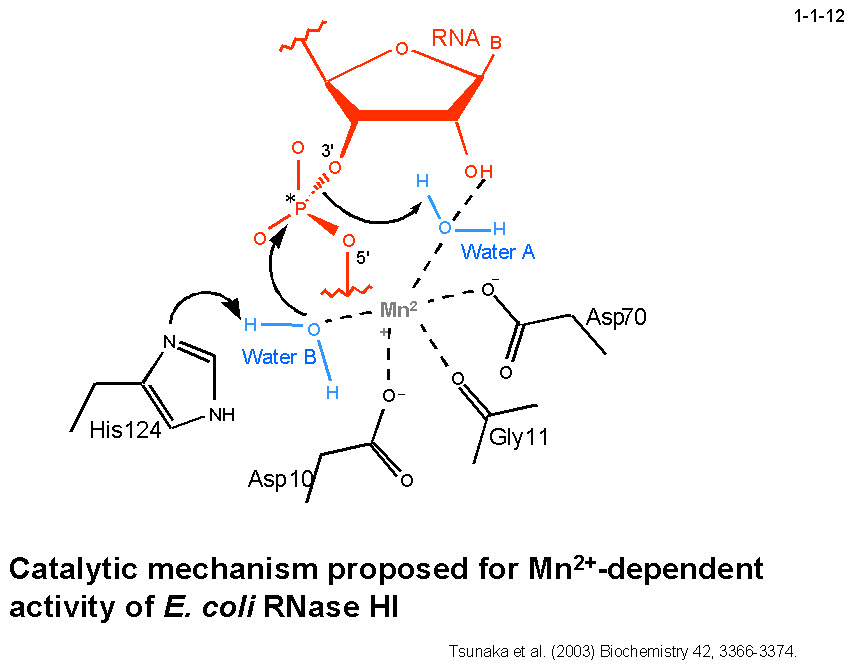
私達はまた、Mg2+が結合した場合とMn2+が結合した場合では大腸菌 RNase HI の触媒機構は異なることを提案している。
つまり、Mg2+が結合した場合は Asp10 と Asp70 が2価金属イオンの結合に、Glu48 と Asp134 が一般酸塩基として働く水分子の保持にそれぞれ必要であるのに対して(1-1-11)、
Mn2+(1つめ)が結合した場合は Glu48 と Asp134 の代わりに Mn2+自身が活性に必要な水分子を保持する(1-1-12)。
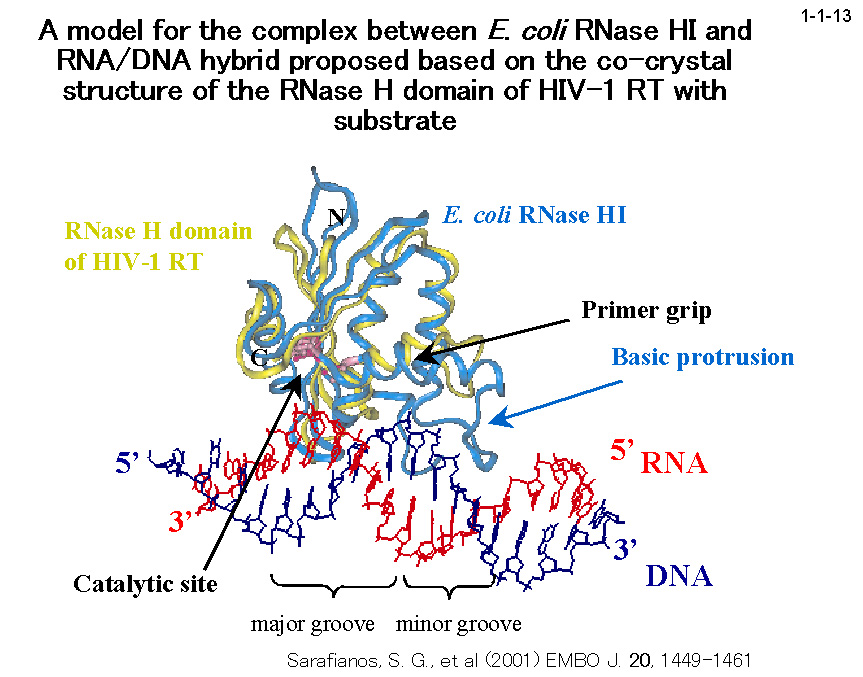
大腸菌 RNase HI の基質認識機構に関しては、HIV-1 逆転写酵素とポリプリントラクト(PPT)を含む RNA/DNA ハイブリドの
共結晶構造を参考にして構築されたモデル(1-1-13)に基づき、以下のような提案を行っている。1)本酵素は基質の小さい溝(minor groove)に結合する、
2)結合に関与するのは本酵素のα2 ヘリックスと基質の DNA 鎖で、本酵素と RNA 鎖は直接結合には関与しない、3)RNA 鎖の位置は DNA 鎖により固定されるので、
RNA/DNA ハイブリドの小さい溝の幅が丁度よい時だけ RNA 鎖は本酵素の活性部位と接触し切断される。つまり、二本鎖 RNA や二本鎖 DNA が本酵素により切断されないのは、
その小さい溝の幅がそれぞれ大きすぎるか小さすぎるために基質が本酵素の活性部位と接触できないためと考えられる。
このような酵素基質複合体の構造モデルを検証するためにはその結晶構造を決定する必要がある。私達は、大腸菌 RNase HI だけでなく、
超好熱菌 RNase HII や好熱菌 RNase HIII を用いて、酵素基質複合体の結晶化を試みている。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}